Механизм наследственности
Das Sein ist ewig; denn Gesetze Bewahren die lebend'gen Schätze, Aus weichen sich das All geschmückt.
1 (Бытие вечно; ибо существуют законы, охраняющие сокровища жизни, которыми украшает себя Вселенная.-Гете.)
Ожидание классического физика, будучи далеко не тривиальным, оказывается неверным
Итак мы пришли к заключению, что организмы со всеми протекающими в них биологическими процессами должны иметь весьма "многоатомную" Структуру, и для них необходимо, чтобы случайные "одноатомные" явления не играли в них слишком большой роли. Существенно, говорит "наивный физик", чтобы организм мог, так сказать, иметь достаточно точные физические законы, на которых он может строить организацию своей исключительно регулярной и хорошо упорядоченной работы. В какой степени приложимы к реальным биологическим фактам эти заключения, достигнутые, говоря биологически, a priori (то есть с чисто физической точки зрения)?
На первый взгляд может показаться, что эти заключения довольно тривиальны. Биолог, скажем, лет 30 назад мог утверждать, что хотя для популярного лектора вполне уместно подчеркнуть значение статистической физики в организме, как и повсюду, однако этот пункт является все же, пожалуй, чересчур избитой истиной. Ибо действительно, не только тело взрослого индивидуума любого высокоразвитого вида, но и каждая клетка его содержит "космическое" число единичных атомов всех родов. И каждый отдельный физиологический процесс, который мы наблюдаем внутри клетки или в ее взаимодействии с внешней средой, кажется - или казалось 30 лет назад, - вовлекает такое огромное количество единичных атомов и единичных атомных процессов, что точное выполнение всех относящихся сюда законов физики и физической химии было бы гарантировано даже при весьма высоких требованиях статистической физики в отношении "больших чисел". Эти требования я только что иллюстрировал правилом √n.
Теперь мы знаем, что такая точка зрения была бы ошибочной. Как мы сейчас увидим, невероятно маленькие группы атомов, слишком малые, чтобы они могли проявлять точные статистические законы, играют главенствующую роль в весьма упорядоченных и закономерных явлениях внутри живого организма. Они управляют видимыми признаками большого масштаба, которые организм приобретает в течение своего развития, они определяют важные особенности его функционирования, и во всем этом выявляются весьма отчетливые и строгие биологические законы.
Я должен начать с краткого подведения итога тому положению, которое имеет место в биологии и, более узко, в генетике; другими словами, я должен суммировать современное состояние знаний в такой области, где я не являюсь авторитетом. Этого нельзя избежать, и поэтому я извиняюсь, особенно перед всяким биологом, за дилетантский характер изложения. С другой стороны, я прошу разрешения изложить вам господствующие представления более или менее догматично. От "бедного" физика-теоретика нельзя ожидать, чтобы он сделал что-нибудь, подобное компетентному обзору экспериментальных данных состоящих из большого количества длинных' и великолепно переплетающихся серий экспериментов по скрещиванию, задуманных с беспрецедентным остроумием, с одной стороны, и из прямых наблюдений над живой клеткой, проведенных со всей утонченностью современной микроскопии,- с другой.
Наследственный шифровальный код (хромосомы)
Разрешите мне воспользоваться словом "план" (pattern) организма в том смысле, в котором биолог называет его "планом в четырех измерениях", обозначая этим не только структуру и функционирование организма во взрослом состоянии или на любой другой определенной стадии, но организм в его онтогенетическом развитии, от оплодотворенной яйцевой клетки до стадии зрелости, когда он начинает размножаться. Теперь известно, что весь этот целостный план в четырех измерениях определяется структурой всего одной клетки, а именно - оплодотворенного яйца. Более того, мы знаем, что он в основном определяется структурой только одной небольшой части этой клетки, ее ядром. Такое ядро в обычном "покоящемся состоянии" клетки представляется как сетка хроматина1 распределенного в пузырьке внутри клетки. Но в жизненно важных процессах клеточного деления (митоз и мейоз, см. ниже) видно, что ядро состоит из набора частиц, обычно имеющих форму нитей или палочек и называемых хромосомами, количество которых 8 или 12, или, например у человека, 48. Но в действительности я должен был бы написать эти (взятые для примера) числа, как 2x4, 2 х 6,..., 2 х 24, и говорить о двух наборах, чтобы пользоваться этим выражением в том обычном значении, в каком оно употребляется биологом. Потому что, хотя отдельные хромосомы иногда отчетливо различимы и индивидуализированы по форме и размеру, эти два набора почти полностью подобны друг другу. Как мы скоро увидим, один набор приходит от матери (яйцевая клетка) и один- от отца (оплодотворяющий сперматозоид). Именно эти хромосомы или, возможно, только осевая или скелетная нить того, что мы видим под микроскопом как хромосому, содержат в виде своего рода шифровального кода весь "план" будущего развития индивидуума и его функционирования в зрелом состоянии. Каждый полный набор хромосом содержит весь шифр, так что имеются, как правило, две копии последнего в оплодотворенной яйцевой клетке, которая представляет самую раннюю стадию будущего индивидуума.
1 (Это оленю значит "вещество, которое окрашивается" в определенном процессе окрашивания, применяемом в микроскопической технике. )
Называя структуру хромосомных нитей шифровальным кодом, мы разумеем, что всеохватывающий ум, вроде такого, который некогда представлял себе Лаплас и которому каждая причинная связь была бы непосредственно открыта, мог бы, исходя из структуры хромосом, сказать, разовьется ли яйцо при благоприятных условиях в черного петуха или в крапчатую курицу, в муху или в растение маиса, в рододендрон, жука, мышь или человека. К этому мы можем прибавить, что внешность различных яйцевых клеток очень часто бывает замечательно сходной, и даже когда это не так (как в случае огромных яиц птиц и рептилий), то все же различие оказывается не столько в существенных структурах, сколько в том питательном материале, который в этих случаях добавляется по понятным причинам.
Но термин шифровальный код, конечно, слишком узок. Хромосомные структуры служат в то же время и инструментом, осуществляющим развитие, которое они же предвещают .Они являются и кодексом законов и исполнительной властью или, употребляя другое сравнение, они являются и планом архитектора и силами строителя в одно и то же время.
Рост тела путем клеточного деления (митоз)
Как хромосомы ведут себя в онтогенезе1?
1 (Онтогенез-это развитие индивидуума в течение его жизни, в противоположность филогенезу - развитию вида в течение геологических периодов.)
Рост организма осуществляется последовательными клеточными делениями. Такое клеточное деление называется митозом. Оно является в жизни клеток не столь частым событием, как этого можно ожидать, учитывая огромное количество клеток, из которых состоит наше тело. Вначале рост идет быстро, и яйцо делится на две "дочерние клетки", которые затем дают поколение из четырех клеток, далее из 8, 16, 32, 64... и т. д. Частота деления не будет оставаться одинаковой во всех частях растущего тела, и это нарушает регулярность этих чисел. Но из их быстрого увеличения можно вывести путем простого вычисления, что в среднем достаточно 50 или 60 последовательных делений, чтобы произвести количество клеток1 , имеющихся у взрослого человека, или, скажем, в десять раз большее количество, принимая во внимание смену клеток в течение жизни. Таким образом, клетки моего тела, в среднем, оказываются только пятидесятыми или шестидесятыми потомками того яйца, которым я когда-то был.
1 (Весьма грубо - сто или тысячу (английских) биллионов.)
В митозе каждая хромосома удваивается
Как ведут себя хромосомы в митозе? Они удваиваются, удваиваются оба набора, обе копии шифра. Этот процесс представляет чрезвычайный интерес и его интенсивно изучали, но он слишком сложен для того, чтобы описывать здесь его детали. Основное заключается в том, что каждая из двух дочерних клеток получает "приданое", состоящее из обоих наборов хромосом, в точности подобных тем, какие были у родительской клетки. Таким путем все телесные клетки совершенно подобны друг другу в отношении их хромосомного сокровища1. Каждая, даже наименее важная отдельная клетка обязательно обладает полной (двойной) копией шифровального кода. Как бы мало мы ни понимали этот механизм, мы не можем, однако, сомневаться, что этот факт должен иметь какое-то важное отношение к жизни организма. Несколько времени назад мы узнали из газет, что во время своей африканской кампании генерал Монтгомери требовал, чтобы каждый отдельный солдат его армии был детально информирован о всех его намерениях. Если это верно (а это могло быть, принимая во внимание высокую интеллигентность и надежность его войск), то мы имеем великолепную аналогию нашему случаю, в котором соответствующий факт, конечно, является буквально верным. Самым удивительным представляется сохранение удвоенного хромосомного набора при всех митотических делениях. То, что это является выдающейся чертой генетического механизма, наиболее разительно подтверждается одним-единственным исключением из этого правила, исключением, которое мы и должны теперь рассмотреть.
1 (Биолог извинит меня за то, что в этом кратком наложении я не рассматриваю исключительного случая мозаиков.)
Редукционное деление (мейоз) и оплодотворение (сингамия)
Очень скоро после начала развития особи одна группа клеток резервируется для образования на поздних стадиях так называемых гамет, то есть спермиев или яйцевых клеток (в зависимости от пола особи), необходимых для размножения индивидуума в зрелости.
"Резервируются" - это значит, что они в это время не служат другим целям и испытывают значительно меньше митотических делений. Происходящее в них исключительное, редукционное деление является тем делением, которым завершается развитие гамет у зрелой особи из этих резервированных клеток. Это деление, как правило, происходит лишь незадолго перед тем, как имеет место сингамия. В мейозе двойной хромосомный набор родительской клетки просто разделяется на два единичных набора, каждый из которых идет в одну из двух дочерних клеток -гамет. Другими словами, митотическое удвоение количества хромосом не имеет места в мейозе, количество их остается постоянным, и таким образом каждая гамета получает только половину, то есть только одну полную копию шифровального кода, а не две, например у человека только 24, а не 2*24=48.
Клетки, имеющие только один хромосомный набор, называются гаплоидными (от греческого àπλοδς, единственный). Таким образом, гаметы гаплоидны, а обычные клетки тела диплоидны (от греческого διπλοδς, двойной). Иногда также встречаются индивидуумы с тремя, четырьмя... или, вообще говоря, с многими хромосомными наборами во всех клетках их тела, и они тогда называются триплоидами, тетраплоидами, ... полиплоидами.
В акте сингамии мужская гамета (сперматозоид) и женская гамета (яйцо) - и та и другая гаплоидные клетки - соединяются, чтобы образовать оплодотворенную яйцевую клетку, которая, следовательно, диплоидна. Один из ее хромосомных наборов приходит от матери и один от отца.
Фотографии на табл. I дают нам некоторое представление о том, как выглядят хромосомы под микроскопом. Я обязан ими д-ру Дарлингтону, в книге которого "Работа с хромосомами"1 интересующийся читатель найдет гораздо больше документов такого же рода и беспрецедентной красоты. На цветной табл. II я пробую дать схематический обзор трех основных процессов - митоза, мейоза и сингамии у маленькой плодовой мушки Drosophila, которая имеет гаплоидное хромосомное число 4 и как объект экспериментирования играет такую выдающуюся роль в современной генетике. Четыре различные хромосомы условно отмечены различным цветом: зеленым, черным, красным и синим. Во вставке (а) хромосомный набор диплоидной клетки тела показан в большом масштабе. Это служит только для пояснения диаграмм (b), (с) и (dj, которые даны в уменьшенном масштабе и чисто схематически. Разрешите мне очистить свою совесть признанием, что в случае мейоза я, как в предыдущем описании, так и в диаграмме, допустил некоторое упрощение, которое, однако, для нашей цели совершенно несущественно2
1 (С.D.Darlington, The Handling of Chromosomes. Allen and Unwin, 1942.)
2 (В действительности мейоз является не одним делением без удвоения числа хромосом, а двумя делениями, почти сливающимися одно с другим, но только с одним удвоением. Результат же просто тот, что в одно и то же время образуются не две гаплоидные гаметы, а четыре.)
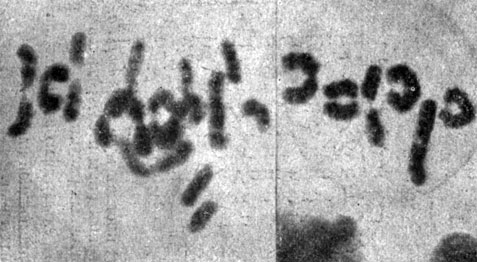
Таблица I. Сблизившиеся попарно хромосомы в материнских клетках пыльцы двух видов Tradescantia. Справа - шесть пар в клетках, фиксированных и окрашенных в ацеторсеине. Слева - двенадцать пар в живой клетке, сфотографированной в ультрафиолетовом свете(* 1000)
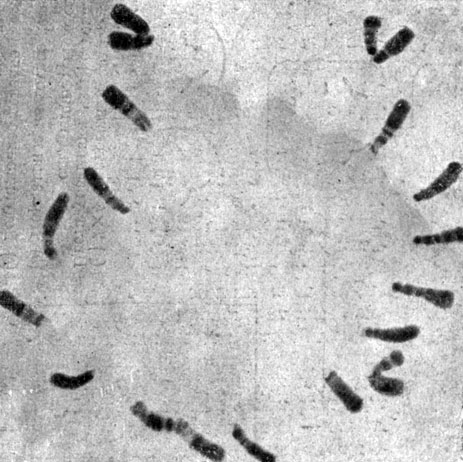
Таблица I. Голодающие хромосомы1 в пыльцевых зернах Fritillaria pudica, полученные путем воздействия холодом. Бледные полосы - инертные сегменты (X 1800)
1 (Здесь разумеется нуклеиповокислое голодание, применяемое с целью получения более совершенно окрашивающихся препаратов.-Прим. пер.)
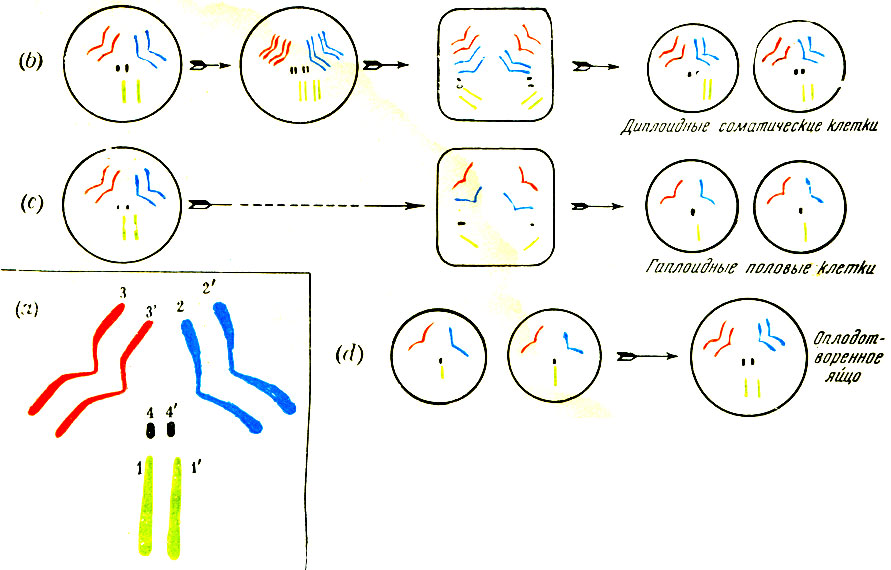
Таблица II
(а) 2*4 хромосомы в соматических клетках мушки Drosophila. Цвета указывают четыре гомологичных пары и соблюдаются в следующих весьма схематичных рисунках.
(b) Нормальное клеточное деление(митоз) диплоидной клетки.
(с) Редукционное деление(мейоз) диплоидной клетки и образование гаплоидных гамет.
(d) Оплодотворение(сингамия). Гаплоидные мужская и женская гаметы соединяются, чтобы образовать диплоидное оплодотворенное яйцо.
Гаплоидные особи
Оговорки требует еще один пункт. Хотя Гаплоидные он и не является существенным для нашей особи цели, он, однако, действительно интересен, поскольку показывает, что в каждом единичном наборе хромосом содержится совершенно полный шифровальный код всего "плана" организма.
Имеются примеры мейоза, за которым оплодотворение следует не сразу, и гаплоидная клетка ("гамета") подвергается в это время большому количеству митотических клеточных делений, в результате чего возникает целая гаплоидная особь. Это случай самцов пчелы - трутней, которые развиваются партеногенетически, то есть из неоплодотворенных и поэтому гаплоидных яиц царицы. Трутень не имеет отца! Все клетки его тела гаплоидны.
Если хотите, можно назвать его гигантски увеличившимся сперматозоидом и, действительно, известно, что функционировать в качестве такового является его единственной жизненной задачей. Однако, может быть, это не серьезная точка зрения. Ибо этот случай не является единичным. Есть семейства растений, где гаплоидные клетки, которые образуются при мейозе и называются спорами, падают на землю как семена и развиваются в настоящие гаплоидные растения, сравнимые по размеру с диплоидными. На рис. 5 изображен грубый набросок мха, хорошо известного в наших лесах.
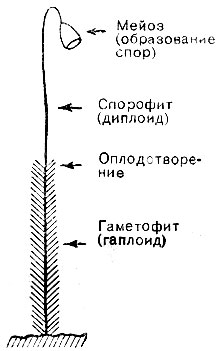
Рис.5. Чередование поколений
Покрытая листочками нижняя часть представляет собой гаплоидное растение, называемое гаметофитом, потому что в своем верхнем конце оно развивает половые органы и гаметы, которые путем оплодотворения производят обычное диплоидное растение - голый стебель с семенной коробочкой на верхушке. Эта часть растения называется спорофитом, так как путем мейоза она производит споры, находящиеся в капсуле на верхушке. Когда капсула открывается, споры падают на землю и развиваются в облиственный стебель. Этот процесс метко назван чередованием поколений. Вы можете, если хотите, рассматривать обычный случай человека и животных с той же точки зрения. Но "гаметофитом" здесь является, как правило, весьма коротко живущее одноклеточное поколение, сперматозоид или яйцевая клетка. Наше тело соответствует спорофиту. Наши "споры" - это резервные клетки, из которых путем мейоза возникает одноклеточное поколение.
Выдающееся значение редукционного деления
Важным и действительно определяющим судьбу событием в процессе воспроизведения индивидуума является не оплодотворение, а мейоз. Один набор хромосом происходит от отца, один - от матери. Ни случайность, ни судьба не могут помешать этому. Каждый человек1 получает ровно половину своей наследственности от матери и половину от отца. То, что одна линия кажется часто преобладающей, объясняется другими причинами, к которым мы перейдем позже (пол сам по себе, конечно, тоже представляет простейший пример такого преобладания).
1 (Во всяком случае, каждая женщина. Чтобы избежать многословия, я исключил из этого обзора чрезвычайно интересную область определения пола и сцепленных с полом признаков (как, например, так называемая цветная слепота).)
Но когда вы проследите происхождение вашей наследственности вплоть до ваших дедов и бабок, то дело оказывается иным. Разрешите мне обратить ваше внимание на набор хромосом, пришедших ко мне от отца, в частности на одну из них, скажем, на хромосому № 5. Это будет точная копия или того № 5, который мой отец получил от своего отца, или того № 5, который он получил от своей матери. Исход дела был решен (с вероятностью 50: 50 шансов) в мейозе, происшедшем в теле моего отца в ноябре 1886 г. и произведшем тот сперматозоид, который немногими днями позже оказался причиной моего зарождения. Точно та же история могла бы быть повторена относительно хромосом № 1, 2, 3... 24 моего отцовского набора и mutatis mutandis относительно каждой из моих материнских хромосом.
Более того, все 48 результатов являются совершенно независимыми. Даже если бы было известно, что моя отцовская хромосома №5 пришла от моего деда Джозефа Шредингера, для № 7 еще оставались бы равные шансы, что она произошла или от него же, или от его жены Марии, урожденной Богнер.
Кроссинговер. Локализация свойств
Но роль случайности в смешении дедушкиной и бабушкиной наследственности у потомков еще больше, чем это могло показаться из предыдущего описания, в котором молчаливо предполагалось или даже прямо утверждалось, что определенные хромосомы пришли как целое или от бабушки, или от дедушки, другими словами, что единичные хромосомы пришли неразделенными. В действительности это не так или не всегда так. Перед тем как разойтись в редукционном делении, скажем в том, которое происходило в отцовском теле, каждые две "гомологичные" хромосомы приходят в тесный контакт одна с другой и иногда обмениваются друг с другом значительными своими частями таким образом, как это показано на рис. 6 (табл. III дает микрофотографию, на которой виден даже еще более тесный и множественный контакт, чем на схеме). Путем такого процесса, называемого "кроссинговер" (перекрест), два свойства, расположенные в соответственных частях этой хромосомы, будут разделены у внука, который окажется похожим одним из этих свойств на дедушку, а другим на бабушку1.
1 (Автор здесь выражается неточно, говоря о расположении в хромосоме "свойств" или "признаков". Как он сам далее указывает, в хромосоме расположены не сами свойства, а лишь определенные материальные структуры (гены), различия в которых приводят к видоизменениям определенных свойств всего организма в целом. Это надо постоянно иметь в виду, ибо Шредингер все время пользуется кратким выражением "свойства". (Прим. перев.))
Явление кроссинговера, будучи не слишком редким, но и не слишком частым, обеспечивает нас ценнейшей информацией о расположении свойств в хромосомах. Чтобы рассмотреть вопрос полностью, мы должны были бы использовать некоторые представления, которые будут даны только в следующей главе (например, гетерозиготность, доминантность и т.д.), но так как это увело бы нас за пределы размеров этой маленькой книги, разрешите мне просто указать на самый важный пункт.
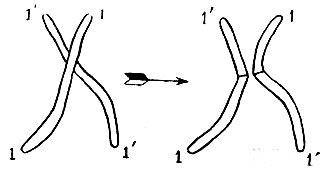
Рис.6. Кроссинговер. Слева - две гомологичные хромосомы в контакте; справа - после обмена и разделения.
Если бы не было кроссинговера, то два признака, за которые ответственна одна и та же хромосома, приходили бы к потомку всегда вместе, и ни одна особь не могла бы получить один из них, не получив также и другого. Два же свойства, определяемые двумя различными хромосомами, либо имели бы шансы 50 : 50 оказаться отделенными друг от друга, либо всегда расходились бы в потомстве к разным особям, а именно тогда, когда эти свойства расположены у предка в гомологичных хромосомах, которые никогда во время мейоза не идут вместе.
Эти правила и отношения нарушаются кроссинговером, вероятность которого может быть установлена путем тщательного регистрирования процента различных комбинаций признаков у потомства в широких экспериментах по скрещиванию, поставленных надлежащим образом для этой цели. Анализируя результаты таких скрещиваний, принимают убедительную рабочую гипотезу, что "сцепление" между двумя свойствами, расположенными в одной хромосоме, тем реже нарушается кроссинговером, чем ближе эти свойства лежат одно к другому. Ибо тогда менее вероятно, что точка разрыва ляжет между ними, тогда как особенности, расположенные ближе к противоположным концам хромосомы, будут разделяться каждым кроссинговером. (То же самое применимо и к объединению в одной хромосоме двух признаков, расположенных ранее в гомологичных хромосомах одного и того же предка.) Таким путем можно ожидать получения из "статистики сцепления" своего рода "карты признаков" внутри каждой хромосомы.
Это ожидание целиком подтвердилось. В случаях, когда была проведена тщательная проверка (главным образом у Drosophila, хотя и не только у нее), оказалось, что изученные признаки действительно распадаются на такое количество отдельных групп, между которыми нет сцепления, сколько имеется различных хромосом (четыре у Drosoyliila). В пределах каждой группы может быть вычерчена линейная карта признаков, количественно выражающая степень сцепления между каждой парой признаков этой группы; поэтому не может быть больших сомнений, что они действительно расположены в хромосоме и притом линейно, как это заставляет думать и самая палочкообразная форма хромосом.
Конечно, схема наследственного механизма, как она описана здесь, еще пуста и бесцветна, даже слегка наивна. Ибо мы не сказали, что следует разуметь под признаком. Рассекать на дискретные "признаки" организм, который является в сущности единым, "целым", представляется неправильным и невозможным. В действительности мы утверждаем в каждом отдельном случае только, что пара предков различалась в определенном, хорошо выраженном отношении (скажем, один имел голубые глаза, а другой - карие) и что потомство сходно в этом отношении или с одним или с другим предком. В хромосоме же мы локализуем место этого различия. (Мы называем его на техническом языке "локус"- или, если мы думаем о гипотетической материальной структуре, которая образует его основу,-"ген".) На мой взгляд, основным представлением служит скорее различие признаков, чем признак сам по себе, несмотря на кажущееся словесное и логическое противоречие в этом утверждении. Различие признаков действительно дискретно, как это выявится в следующей главе, когда мы будем говорить о мутациях и когда представленная выше сухая схема, я надеюсь, приобретет больше жизни и красок.
Максимальный размер гена
Мы только что ввели термин ген для гипотетического материального носителя определенной наследственной особенности. Мы должны теперь подчеркнуть два момента, которые будут иметь большое значение для нашего исследования. Первый момент - это размер, или, лучше сказать, максимальный размер этого носителя; другими словами, - до сколь малого объема можем мы проследить локализацию наследственных потенций. Второй момент - это устойчивость гена выведенная из постоянства "наследственного плана".
В отношении размера имеются два совершенно независимых способа определения. Один основан на генетических данных (эксперименты по скрещиванию), другой - на цитологических данных (прямое микроскопическое наблюдение). Первый способ принципиально достаточно прост. Установив описанным выше путем расположение значительного числа различных признаков (большего масштаба) внутри определенной хромосомы (скажем, у мушки Drosophila), мы, чтобы получить требуемую величину, должны только разделить измеренную длину этой хромосомы на количество признаков и умножить на поперечное сечение. Ибо, конечно, мы рассматриваем как отдельные признаки только такие, которые иногда разделяются кроссинговером и не могут быть обусловлены одной и той же (микроскопической или молекулярной) структурой. С другой стороны, ясно, что наш расчет может дать только максимальный размер, потому что количество признаков, изолированных генетическим анализом, непрерывно растет по мере того, как работа идет вперед.
Другая оценка размера, хотя и основанная на микроскопическом наблюдении, в действительности является гораздо менее прямой. Определенные клетки Drosophila (именно, клетки слюнных желез) оказываются по каким-то причинам гигантски увеличенными, и это касается и их хромосом. В этих последних вы различаете скученный рисунок из поперечных темных полосок, пересекающих нить (табл. IV). Дарлингтон подметил, что число этих полосок (2000 в рассматриваемом случае), хотя и заметно больше, но все же того же самого порядка, как и число генов, локализованных в той же самой хромосоме на основании экспериментов по скрещиванию. Он склонен рассматривать эти полоски как действительные гены (или границы между генами). Разделив длину хромосомы, измеренную в клетке нормального размера, на число полосок (2000), он находит объем гена равным кубу со стороной в 300 А ?. Учитывая всю грубость расчетов, мы можем считать, что такой же размер получается и первым методом.
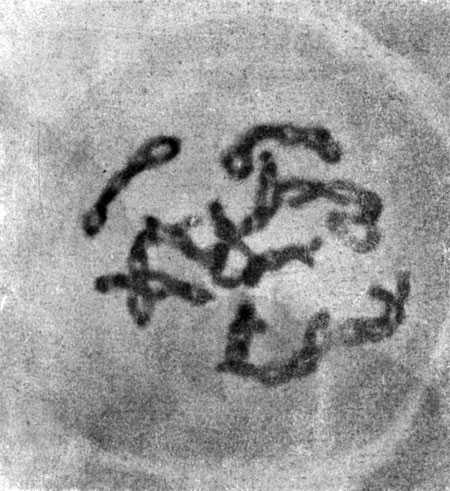
Таблица III. Двенадцать пар сблизившихся хромосом в материнских клетках пыльцы Fritillaria chitralensis. Точки пересечения петель показывают места кроссинговера между партнерами (X 1600)
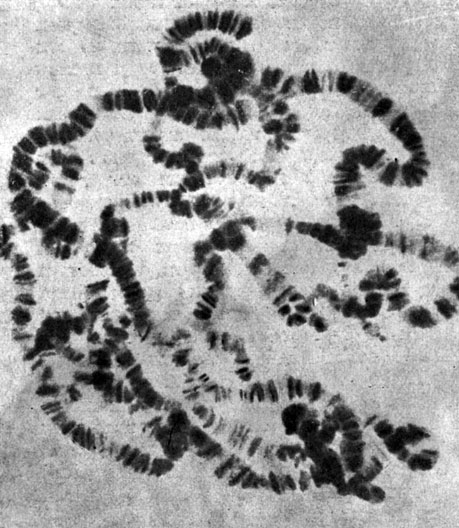
Таблица IV. Покоящееся ядро клетки слюнной железы мушки Drosophila melanogaster. Гены прошли восемь циклов удвоения и поэтому выглядят как серии поперечных полосок, из которых каждая содержит 256 генов. Более крупные гены дают сильнее окрашенные полоски (X 1500)
Малые числа
Подробное обсуждение отношения статистической физики ко всем фактам, которые я изложил (или я, может быть, должен сказать - отношение этих фактов к применению статистической физики в живой клетке), последует позже. Но разрешите мне привлечь ваше внимание сейчас к тому обстоятельству, что 300( А) ? - это только около 100 или 150 атомных расстояний в жидкости или в твердом теле, так что ген, несомненно, содержит не более миллиона или нескольких миллионов атомов. Согласно статистической физике, а это значит- согласно физике вообще, такое число слишком мало (с точки зрения ?n чтобы обусловить упорядоченное и закономерное поведение. Оно было бы слишком мало, даже если бы все эти атомы исполняли одинаковую роль, как в газе или капле жидкости, а ген, почти несомненно, как раз не является гомогенной каплей жидкости. Он, вероятно, большая протеиновая молекула, где каждый атом, каждый радикал, каждое гетероциклическое кольцо играет индивидуальную роль, более или менее отличную от роли любых сходных атомов, радикалов или колец. Это, во всяком случае, точка зрения таких ведущих генетиков нашего времени, как Холдэн и Дарлингтон, и мы скоро должны будем обратиться к генетическим экспериментам, которые почти доказывают это.
Постоянство
Обратимся теперь ко второму, весьма важному вопросу: с какой степенью постоянства мы сталкиваемся в наследственных особенностях и что мы поэтому должны приписать тем материальным структурам, которые их несут.
Ответ на это может быть дан без какого-либо специального исследования. Простой факт, что мы говорим о наследственных особенностях, указывает, что мы признаем это постоянство почти абсолютным. Ибо мы не должны забывать, что от родителя к ребенку передается вовсе не отдельная особенность: орлиный нос, короткие пальцы, предрасположение к ревматизму, гемофилия, дихромазия и т. д. Такие черты удобно вычленять для изучения законов наследственности. Но в действительности из поколения в поколение, без заметного изменения в течение столетий - хотя и не в течение десятков тысяч лет,- передается весь (четырехмерный) план "фенотипа", вся видимая природа индивидуума. При этом в каждом поколении передача осуществляется материальной структурой ядер тех двух клеток, которые соединяются при оплодотворении. Это- "чудо"; имеется только одно еще большее "чудо", хотя и связанное тесно с первым, но относящееся уже к другой сфере. Я подразумеваю тот факт, что мы, чье существование целиком основано на удивительной игре именно этого механизма наследственности, все же обладаем способностью узнать о нем так много. Мне представляется, что в отношении первого чуда наши знания могут дойти едва ли не до полного понимания. Что касается второго, то возможно, что оно вообще лежит за пределами человеческого познания.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить ссылку на страницу источник:
http://physiclib.ru/ 'Библиотека по физике'