Мутации
Und was in schwankender Erscheinung schwebt, Befestiget mit dauernden Gedanken.
1 (И то, что носится в колеблющихся очертаниях, закрепляется в прочных мыслях. - Гете.)
"Скачкообразные" мутации - поле действия естественного отбора
Основные факты, которые мы только что выдвинули в доказательство устойчивости, приписываемой генной структуре, может быть, хорошо известны нам и не покажутся поразительными и убедительными. Но на этот раз поговорка, что исключения подтверждают правило, действительно верна. Если бы не было исключений в сходстве между детьми и родителями, мы были бы лишены не только всех прекрасных экспериментов, открывших нам механизм наследственности, но также и грандиозного, миллионнократного эксперимента природы, кующего виды путем естественного отбора и выживания наиболее приспособленных.
Разрешите мне взять последнюю важную проблему исходной точкой для того, чтобы представить относящиеся сюда факты, опять с извинением и с напоминанием, что я не биолог.
Мы теперь определенно знаем, что Дарвин ошибался, когда считал, что материалом, на основе которого действует естественный отбор, служат малые, непрерывные, случайные изменения, обязательно встречающиеся даже в наиболее однородной популяции. Потому что было доказано, что эти изменения не наследственны. Этот факт достаточно важен, чтобы его кратко проиллюстрировать. Если вы возьмете урожай чистосортного ячменя и измерите у каждого колоса длину остей, а затем вычертите результат вашей статистики, вы получите колоколообразную кривую, как это показано на рис. 7.
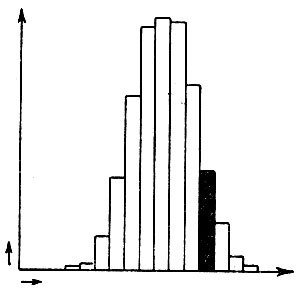
Рис. 7- Статистика длины остей в чистосортном ячмене. Черная группа должна быть отобрана для посева (детали взяты не из реального эксперимента, но подобраны только для иллюстрации)
На этом рисунке количество колосьев с определенной длиной остей отложено против соответствующей длины остей. Другими словами, преобладает известная средняя длина остей, а отклонения в том и другом направлении встречаются с определенными частотами. Теперь выберите группу колосьев, обозначенную черным, с остями, заметно превосходящими среднюю длину, но группу достаточно многочисленную, чтобы при посеве в поле она дала новый урожай. Проделывая подобный же статистический опыт, Дарвин ожидал бы, что для нового урожая кривая сдвинется вправо. Другими словами, он ожидал бы, что отбор произведет увеличение средней величины остей. Однако на деле этого не случится, если использовался действительно чистосортный ячмень. Новая статистическая кривая, полученная для отобранного урожая, будет вполне подобна первой, и то же самое случилось бы, если бы были отобраны для посева колосья с особенно короткими остями.
Отбор не дает результата, потому что малые, непрерывные различия не наследуются. Они, очевидно, не обусловлены строением наследственного вещества, они случайны. Но около 40 лет тому назад голландец де-Фриз открыл, что в потомстве даже совершенно чистосортных линий появляется очень небольшое число особей - скажем, две или три на десятки тысяч - с малыми, но "скачкообразными" изменениями. Выражение "скачкообразные" означает здесь не то, что изменения очень значительны, а только факт прерывистости, так как между неизмененными особями и немногими измененными нет промежуточных форм. Де-Фриз назвал это мутацией. Существенной чертой тут является именно прерывистость. Физику она напоминает квантовую теорию - там тоже не наблюдается промежуточных ступеней между двумя соседними энергетическими уровнями. Физик был бы склонен мутационную теорию де-Фриза фигурально назвать квантовой теорией биологии. Мы увидим позже, что это значительно больше, чем фигуральное выражение. Мутации действительно обязаны своим происхождением квантовым скачкам в генной молекуле. Но квантовой теории было еще только два года от роду, когда де-Фриз впервые опубликовал свое открытие в 1902 г. Неудивительно, что потребовалось целое поколение, чтобы установить связь между ними!
Они действительно размножаются, т.е. они наследуются в совершенстве
Мутации наследуются так же хорошо, как первоначальные неизмененные признаки. Например, в первом урожае ячменя, рассмотренном выше, могло оказаться несколько колосьев с размером остей, далеко выходящим за пределы изменчивости, показанной на рис. 7, скажем, совсем без остей. Они могли представлять де-фризовскую мутацию и стали бы поэтому размножаться действительно в совершенстве, то есть все их потомки были бы также без остей.
Следовательно, мутация определенно является изменением в наследственном багаже и должна обусловливаться каким-то изменением наследственной субстанции. В самом деле, большинство важных экспериментов, открывших нам механизм наследственности, состояло в тщательном анализе потомства, полученного путем скрещивания мутировавших (а во многих случаях даже множественно мутировавших) индивидуумов с немутировавшими или с иначе мутировавшими. С другой стороны, в силу их свойства действительно передаваться потомкам, мутации служат также подходящим материалом и для естественного отбора, который может работать над ними и производить виды, как это описано Дарвином, элиминируя неприспособленных и сохраняя наиболее приспособленных.
В дарвиновской теории нужно только заменить его "небольшие случайные вариации" мутациями (совсем как в квантовой теории "квантовый скачок" заменяет собой "непрерывные переходы энергии"). Во всех других отношениях в теории Дарвина оказались необходимыми лишь очень небольшие изменения, во всяком случае если я правильно понимаю точку зрения, которой придерживается большинство биологов1.
1 (Широко обсуждался вопрос о том, не помогает ли естественному отбору (если не заменяет его) отмечавшаяся тенденция появления полезных или выгодных мутаций. Моя личная точка зрения по этому вопросу не имеет значения. Но необходимо оговорить, что возможность "направленных мутаций" не принимается во внимание в дальнейшем изложении. Более того, я не могу входить здесь также в обсуждение взаимодействия генов-"модификаторов" и "полимерных" генов, какими бы важными ни были эти вопросы для действительного механизма отбора и эволюции.)
Локализация. Рецессивность и доминантность
Теперь мы должны рассмотреть некоторые другие важнейшие факты и представления, касающиеся мутаций, - опять в несколько догматической форме, не показывая, как эти факты и представления возникли один за другим из экспериментальных данных.
Мы должны были бы ожидать, что определенная мутация вызывается изменением в определенной области одной из хромосом. И так это и есть. Важно констатировать: мы твердо знаем, что это изменение происходит только в одной хромосоме и не возникает одновременно в соответствующем "локусе" гомологичной хромосомы. Схематически это показано на рис. 8, где крестом отмечен мутировавший локус.
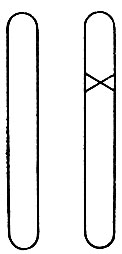
Рис.8. Гетерозиготный мутант. Крестом отмечен мутировавший ген
Факт, что затронута только одна хромосома, обнаруживается, когда мутировавшая особь (часто называемая "мутант") скрещивается с немутировавшей. Ибо при этом ровно половина потомства обнаруживает мутантный признак, а половина - нормальный. Это и есть именно то, чего следует ожидать в результате расхождения у мутанта двух хромосом в мейозе и что показано весьма схематично на рис. 9.
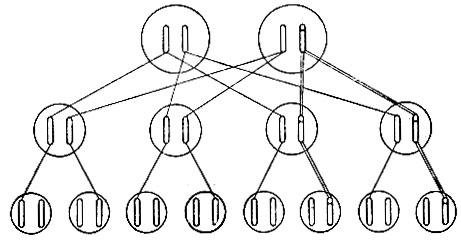
Рис. 9. Наследование мутации. Прямые линии указывают передачу нормальной хромосомы, а двойные - передачу мутировавшей хромосомы. Хромосомы третьего поколения, происхождение которых не указано, приходят невключенными в схему супругов особей второго поколения. Предполагается, что эти супруги не родственны и свободны от мутаций
На этом рисунке дана родословная, где каждый индивидуум (трех последовательных поколений) представлен просто парой хромосом. Пожалуйста, учтите, что если бы обе хромосомы мутанта были изменены, то все дети имели бы одну и ту же (смешанную) наследственность, отличную от наследственности каждого родителя.
Но экспериментировать в этой области не так просто, как могло показаться из вышесказанного. Дело усложняется вторым важным обстоятельством, а именно тем, что мутации весьма часто бывают скрытыми. Что это значит?
У мутантной особи две "копии шифровального кода" больше уже не одинаковы; они представляют два различных "толкования" или две "версии", во всяком случае в том месте, где произошла мутация. Может быть, полезно указать сразу, что хотя это и соблазнительно, но было бы совершенно неверно рассматривать первоначальную версию как "ортодоксальную", а мутантную версию как "еретическую". Мы должны рассматривать их в принципе как равноправные, ибо и нормальные признаки в свое время также возникли путем мутаций.
Действительно, признаки мутантного индивидуума, как общее правило, соответствуют или той или другой версии, причем эта версия может быть как нормальной, так и мутантной. Версия, которой следует особь, называется доминантной, противоположная-рецессивной; другими словами, мутация называется доминантной или рецессивной в зависимости от того, проявляет ли она свой эффект сразу или нет.
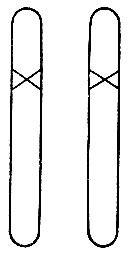
Рис.10. Гомозиготный мутант, полученный в одной четверти потомства при оплодотворении гетерозиготных мутантов (см. рис.8) или при скрещивании их между собой
Рецессивные мутации даже более часты, чем доминантные, и бывают весьма важными, хотя они не сразу обнаруживаются. Чтобы л изменить свойства организма, они должны присутствовать в обеих хромосомах (рис. 10). Такие индивидуумы могут быть получены, когда два одинаковых рецессивных мутанта скрещиваются между собой или когда мутант скрещивается сам с собой. Последнее возможно у гермафродитных растений и происходит даже самопроизвольно. Простое рассуждение показывает, что в этих случаях около четверти потомства будет мутантной внешности.
Введение некоторых технических терминов
Для большей ясности здесь следует объяснить некоторые технические термины. То, что я называю "версией шифровального кода" - будь она первоначальной или мутантной, - принято обозначать термином "аллель". Когда версии различны, как это показано на рис. 8, особь называется гетерозиготной в отношении этого локуса. Когда они одинаковы, как, например, в немутировавших особях или в случае, изображенном на рис. 10, они называются гомозиготными. Таким образом, рецессивные аллели влияют на признаки только в гомозиготном состоянии, тогда как доминантные аллели производят один и тот же признак как в гомозиготном, так и в гетерозиготном состоянии.
Цвет очень часто доминирует над отсутствием цвета (или белизной). Так, например, горох будет цвести белым цветом, только когда он имеет "рецессивную аллель, ответственную за белый цвет" в обеих соответствующих хромосомах, то есть когда он "гомозиготен по белому"; он будет тогда давать чистое потомство, и все его потомки будут белыми. Но уже одна "красная аллель" (в то время как другая белая - "гетерозиготная особь") сделает цветок красным, и совершенно таким же сделают его и две красные аллели ("гомозиготная особь"). Различие последних двух случаев станет выявляться только в потомстве, когда гетерозиготные красные будут производить некоторое количество белых потомков, а гомозиготные красные будут давать чистое потомство.
То, что две особи могут быть совершенно подобны по внешности и, однако, различаться наследственно, столь важно, что желательно дать этому точную формулировку. Генетик говорит, что у особей один и тот же фенотип, но различный генотип. Содержание предыдущих параграфов может быть, таким образом, суммировано в кратком, но высоко техническом выражении: рецессивная аллель влияет на фенотип, только когда генотип гомозиготен.
Мы будем прибегать время от времени к этим техническим выражениям, напоминая читателю их значение, когда это необходимо.
Вредное действие родственного скрещивания
Рецессивные мутации, пока они гетерозиготны, не служат, конечно, материалом для естественного отбора. Если они вредны, как это часто бывает с мутациями, они, тем не менее, не отбрасываются, потому что они скрыты.
Отсюда следует, что очень большое количество неблагоприятных мутаций может накопляться и не причинять непосредственного вреда. Но они, конечно, передаются половине потомства, и это применимо как к человеку, так и к скоту, домашней птице и другим видам, хорошие физические качества которых имеют для нас непосредственное значение. На рис. 9 предполагается, что мужской индивидуум (скажем, для конкретности, я сам) несет такую рецессивную вредную мутацию в гетерозиготном состоянии, так что она не проявляется. Предположим, что моя жена не имеет ее. Тогда половина наших детей (второй ряд) будет также нести ее, и притом опять в гетерозиготном состоянии. Если все они вступят в брак с немутантными партнерами (опущенными в диаграмме, чтобы избежать путаницы), четвертая часть наших внуков в среднем будет затронута подобным же образом.
Никакой опасности вредных проявлений не возникнет до тех пор, пока такие затронутые индивидуумы не поженятся между собой. Тогда, как показывает простой расчет, четвертая часть детей окажется гомозиготной и проявит вредную мутацию. За исключением самооплодотворения (возможного только у гермафродитных растений) наибольшую опасность представлял бы брак между моим сыном и моей дочерью. Каждый из них имеет одинаковые шансы быть в скрытом виде затронутым или незатронутым, и потому одна четвертая часть таких кровосмесительных союзов была бы опасна, поскольку четвертая часть детей от такого брака проявляла бы вредный признак. Размер опасности для каждого отдельного ребенка, рожденного при кровосмешении, равен, таким образом, 1:16.
Сходные рассуждения показывают, что размер опасности составляет 1:64 для потомства в случае брака моих внуков, которые в то же время являются двоюродными братом и сестрой. Это уже не кажется таким страшным, и действительно, последний случай брака обыкновенно считается терпимым. Но не надо забывать, что мы анализировали последствия только одного скрытого повреждения у одного партнера из пары предков ("я и моя жена"). В действительности же оба они, весьма возможно, несут в себе более чем один латентный недостаток этого рода. Если вы знаете, что вы сами носите определенный скрытый недостаток, вы должны предполагать с вероятностью 1:8, что ваши двоюродные братья и сестры также разделяют его с вами!
Эксперименты с растениями и животными, по-видимому, указывают, что в добавление к сравнительно редким дефектам серьезного характера имеется масса меньших, случайные комбинации которых ухудшают в целом потомство от родственных скрещиваний. Поскольку мы более не склонны удалять неудачных потомков тем жестоким путем, каким пользовались лакедемоняне на Тайгетской скале1, мы должны обращать особенно серьезное внимание на близкородственные браки у человека, для которого естественный отбор наиболее приспособленных большей частью ограничен, и даже более того, - обращен в свою противоположность. Антиселективное действие современных массовых убийств здоровых юношей всех национальностей вряд ли оправдывается соображениями, что в более первобытных условиях война могла иметь положительную ценность для отбора, давая возможность пережить наиболее приспособленным племенам
1 (Слабых детей сбрасывали со скалы и таким образом старались освободить свой народ от слабых и больных потомков. (Прим. перев.))
Общие и исторические замечания
Представляется удивительным, что рецессивные аллели в гетерозиготном состоянии полностью подавляются доминантными и совершенно не производят видимого действия. Надо, по крайней мере, упомянуть, что из этого имеются исключения. Когда гомозиготный белый львиный зев скрещивается с гомозиготным же малиновым львиным зевом, все непосредственные потомки оказываются промежуточными по окраске, то есть розовыми (а не малиновыми, как можно было ожидать). Более важный случай двух аллелей, выявляющих свое действие одновременно, наблюдается в кровяных группах, но мы не можем вдаваться здесь в это подробнее. Я не был бы удивлен, если бы в конце концов оказалось, что рецессивность может быть различных степеней и что ее обнаружение зависит от чувствительности приемов, применяемых при изучении "фенотипа".
Здесь, может быть, уместно рассказать о ранней истории генетики. Костяком теории, а именно законами передачи в последующие поколения признаков, которыми различались родители, и в частности, открытием рецессивных и доминантных признаков мы обязаны всемирно известному августинскому аббату Грегору Менделю (1822-1884). Мендель ничего не знал относительно мутаций и хромосом. В своем монастырском саду в Брюнне (Брно) он ставил опыты с садовым горошком, культивируя различные сорта, скрещивая их и наблюдая их потомство в 1-м, 2-м, 3-м..., поколениях. Вы можете сказать, что он экспериментировал с мутантами, найдя их уже готовыми в природе. Результаты он опубликовал еще в 1866 г. в трудах "Nalurforschender Verein in Вrunn". Никто, казалось, не интересовался занятиями аббата и никто, конечно, не имел ни малейшего представления о том, что в двадцатом столетии его открытие станет путеводной звездой совершенно новой ветви науки, возможно, наиболее интересной в наши дни. Его работа была забыта, и ее снова обнаружили только в 1900 г. одновременно и независимо друг от друга Корренс (Берлин), де-Фриз (Лейден) и Чермак (Вена).
Необходимость того, чтобы мутации были редким событием
До сих пор мы обращали наше внимание на вредные мутации, которые, может быть, более многочисленны; однако следует определенно указать, что мы встречаемся также и с полезными мутациями. Если самопроизвольная мутация представляет собой небольшую ступеньку в развитии вида, то создается впечатление, что известное изменение "испытывается" вслепую - с риском, что оно может оказаться вредным и в таком случае будет автоматически элиминировано. Отсюда вытекает один очень важный момент. Чтобы быть подходящим материалом для работы естественного отбора, мутации должны быть достаточно редкими событиями, какими они в действительности и оказываются. Если бы они были настолько частыми, что существовала бы большая вероятность появления у одной особи, скажем, дюжины различных мутаций, то вредные, как правило, преобладали бы над полезными, и виды, вместо того чтобы улучшаться путем отбора, оставались бы неулучшенными или погибали бы. Сравнительный консерватизм, являющийся результатом высокой устойчивости генов, имеет очень существенное значение. Аналогию этому можно усмотреть, например, в работе сложного фабричного оборудования на каком-нибудь заводе.
Для развития лучших методов необходимо испытывать различные нововведения, даже непроверенные раньше. Но чтобы выяснить, увеличивают или уменьшают эти нововведения продукцию завода, важно вводить их по одному, тогда как другие части механизма остаются неизмененными.
Мутации, вызванные х-лучами
Мы теперь должны рассмотреть серию чрезвычайно остроумных генетических исследований, которые окажутся наиболее существенными для нашего анализа.
Процент мутаций в потомстве-так называемый темп мутирования - можно увеличить во много раз по сравнению с естественным мутационным темпом, если освещать родителей х - лучами или γ-лучами. Мутации, вызванные таким путем, ничем (за исключением большей частоты) не отличаются от возникающих самопроизвольно, и создается впечатление, что каждая "естественная" мутация может быть тоже вызвана х -лучами. В обширных культурах Drosophila многие особые мутации повторяются снова и снова; они были локализованы в хромосоме, как это описано в § 16, и получили специальные названия. Были обнаружены так называемые "множественные аллели", то есть две или более различных "версий" или "чтений" (в добавление к нормальной немутировавшей) в том же самом месте хромосомного кода. Это означает, что имеются не только два, но три и больше изменений в данном локусе, причем каждые два из них находятся один к другому в отношении "доминантности-рецессивности", когда они оказываются одновременно на своих соответствующих местах в двух гомологичных хромосомах1.
1 (Это не совсем точно. Отмечено, что за исключением "дикого" (обычного) аллеломорфа остальные чаще ведут себя не как доминантные и дают в сочетании друг с другом промежуточные формы. {Прим. перев.))
Эксперименты с мутациями, вызванными х-лучами, создают впечатление, что каждый отдельный "переход", скажем, от нормального инд ивидуума к данному мутанту или наоборот имеет свой индивидуальный "х-лучевой коэффициент", указывающий процент потомства, которое оказывается мутировавшим в данном специальном направлении, если перед зарождением этого потомства родители получили единичную дозу х-лучей.
Первый закон. Мутация является единичным событием
Более того, законы, управляющие частотой появления индуцированных мутаций, крайне просты и бросают чрезвычайно яркий свет на весь вопрос. Я следую здесь изложению Н.В.Тимофеева в Biological Reviews,vol. 9,1934.
В значительной степени оно основывается на собственной прекрасной работе этого автора. Первый закон гласит:
1. Увеличение числа мутаций точно пропорционально дозе лучей, так что можно действительно говорить (как я это и делал) о коэфициенте увеличения.
Мы так привыкли к простой пропорциональности, что склонны недооценивать далеко идущие последствия этого закона. Чтобы оценить их, мы можем вспомнить, что стоимость товара, например, не всегда пропорциональна его количеству. В обычное время то, что вы уже купили шесть апельсинов, может произвести на лавочника такое впечатление, что если вы потом решите взять у него целую дюжину, он, возможно, отдаст вам ее дешевле, чем за двойную цену первых шести. В случае неурожая может случиться обратное. В нашем случае мы заключаем, что первая половина дозы излучения, вызвав, скажем, одну мутацию на тысячу потомков, в то же время совсем не повлияла на остальных потомков ни в сторону предрасположения, ни в сторону иммунизации против мутаций. Ибо в противном случае вторая половина дозы не вызвала бы снова именно одной мутации на тысячу. Мутация, таким образом, не является накопленным результатом последовательного освещения малыми порциями, которые усиливали бы одна другую. Она должна состоять из какого-то единичного явления, происходящего в одной хромосоме во время воздействия х-лучами. Что же это за явление?
Второй закон. Локализация события
На это отвечает второй закон, а именно:
2. Если вы изменяете качество лучей (длину события волны) в широких пределах от мягких х-лучей до довольно жестких γ-лучей, коэфициент остается постоянным при условии, что вы даете ту же самую дозу в так называемых r-единицах. Иначе говоря, коэфициент не изменяется, если вы измеряете дозу общим количеством ионов, возникающих на единицу объема, в подходящем стандартном веществе в течение времени, когда родители подвергаются действию лучей, и в том же самом месте.
В качестве стандартного вещества выбирают воздух, - не только для удобства, но также по той причине, что"ткани организмов состоят из элементов того же среднего атомного веса, как и воздух. Нижний предел числа ионизации или сопровождающих процессов1 (возбуждений) в тканях получается просто умножением количества ионизации в воздухе на отношение плотностей. Таким образом, совершенно ясно (и это подтверждается более детальным исследованием), что единичное явление, вызывающее мутацию, это и есть как раз ионизация (или сходный процесс), происходящая внутри некоторого "критического" объема зародышевой клетки.
1 (Нижний предел, потому что эти другие процессы не учитываются при измерении ионизации, но могут все же иметь значение при вызывании мутаций.)
Каков же этот критический объем? Он может быть установлен из наблюдающейся частоты мутирования путем следующего рассуждения: если при дозе 50 тыс. ионов на 1см3 вероятность мутировать в данном специальном направлении для каждой отдельной гаметы, находящейся в облучаемом пространстве, равна только 1 : 1000, мы заключаем, что критический объем - "мишень", в которую надо "попасть" ионизации, чтобы возникла эта мутация - будет только 1/1000 от 1/50 000 см3, то есть, иначе говоря, одна пятидесятимиллионная см3. Цифры здесь не точны, и я их привел только для иллюстрации. В действительном расчете мы следуем М. Дельбрюку (в совместной работе его, Н. В. Тимофеева и К. Г. Циммера)1. Эта же работа послужит основным источником при изложении теории в следующих двух главах. Дельбрюк приходит к объему только около десяти средних атомных расстояний в кубе и содержащему, таким образом, только 103 атомов. Простейшее истолкование этого результата сводится к тому, что имеется достаточная вероятность возникновения данной мутации, если ионизация (или возбуждение) происходит не далее, чем на расстоянии около "10 атомов в сторону" от определенного места в хромосоме. Более детально мы это обсудим в дальнейшем.
1 (Nachr.a.d.Biologie d.Ges.d.Wiss.Göttingen, Bd.I,s.189,1935.)
Статья Тимофеева содержит практический намек, от упоминания о котором я не могу здесь воздержаться, хотя он, конечно, не имеет отношения к настоящему исследованию. В современной жизни бывает множество случаев, когда человек подвергается действию х-лучей. Прямые опасности, включая такие, как ожог, рак, стерилизация, хорошо известны; сестрам и врачам, постоянно имеющим дело с лучами, обеспечивается специальная защита свинцовыми ширмами, фартуками и т.д. Дело, однако, в том, что даже при успешном отражении этих неизбежных опасностей, грозящих индивиду, существует косвенная опасность возникновения небольших вредных мутаций в зачатковых клетках, мутаций того же рода, как и те, с которыми мы встречались, когда речь шла о неблагоприятных результатах родственного скрещивания. Говоря более выразительно - хотя, возможно, это звучит немного наивно, - вредность брака между двоюродными братом и сестрой может быть очень увеличена тем, что их бабушка в течение долгого времени служила сестрой в рентгеновском кабинете. Это не должно быть поводом для беспокойства отдельного человека. Но всякая возможность постепенного заражения человеческого рода нежелательными скрытыми мутациями должна интересовать человеческое общество.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить ссылку на страницу источник:
http://physiclib.ru/ 'Библиотека по физике'