Фотосинтез
Удостоверившись в том, насколько трудно осуществить процессы, которые позволили бы преобразовать или аккумулировать солнечную энергию, обратимся теперь к вопросу о том, как они протекают в живых клетках. Здесь они осуществляются удивительно экономично. При благоприятных условиях растения могут накапливать до 10% энергии падающей на них солнечной радиации в виде "горючего", молекулы которого имеют чрезвычайно сложное строение. Общее количество энергии, получаемой на земном шаре посредством такого рода преобразований, оценивается по-разному. Считается, что только для своего существования растения расходуют энергии в 10 раз больше, чем в настоящее время потребляет все человечество. Мельчайшие живые организмы, густо населяющие моря, по-видимому, используют еще больше энергии.
Цель проводимых в этой области исследований заключается в том, чтобы научиться воспроизводить хотя бы часть таких процессов и управлять ими. Однако процессы фотосинтеза необычайно сложны и до сих пор в значительной степени остаются непонятными. Их изучение продолжается, а пока особое внимание уделяется применению растений в качестве преобразователей энергии в энергетических системах, созданных человеком и действующих по другому принципу. В этом разделе мы рассмотрим несколько характерных особенностей фотосинтеза растений, чтобы понять возможности их использования в работе энергосистем.
Реакцию фотосинтеза, происходящую в зеленых растениях, часто представляют уравнением
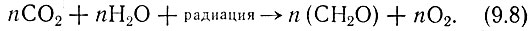
Из него мы видим, что в растениях в результате взаимодействия углекислого газа и воды образуются углеводы и выделяется кислород. Это, конечно, грубое упрощение, которое, однако, позволяет нам уяснить существенные моменты метаболизма растений. Для протекания такого процесса необходимы углекислый газ и вода (первый обычно заимствуется из воздуха, где его содержится около 0,03%). Как показывает уравнение (9.8), на каждую поглощенную молекулу углекислого газа растение выделяет одну молекулу кислорода. Молекулы углеводов участвуют в цепи химических превращении, в результате которых образуются органические вещества, необходимые для роста и развития растении. Простейшей формой углевода n(Cu2O) является глюкоза C6H12O6 (n = 6). В какой-то степени она присутствует в растениях в чистом виде, но чаще всего входит как составная часть в более сложные молекулы жиров, белков и других веществ с высоким содержанием углеводов, щедро представленных в живых растениях. Не останавливаясь на рассмотрении всех этих последовательных преобразований, заметим лишь, что роль катализаторов в них играют очень сложные белковые соединения, называемые ферментами. Энергия, требующаяся для осуществления указанных "строительных" операций, получается при окислении некоторых простейших углеводов с образованием снова углекислого газа CO2 и воды. Эффективность таких операций зависит от содержания в почве микроколичеств определенных элементов. От них иногда зависит, будет ли урожай шедрым или плохим.
Пути движения веществ, участвующих в процессе (9.8), исследовались методом радиоактивных, или "меченых", атомов. Установлено, что кислород выделяется только из воды, но не из углекислого газа. Таким образом, процесс, по существу, сводится к разложению молекул воды, то есть к реакции (9.1), В первых разделах этой главы мы видели, что энергия одного фотона видимого света недостаточна для осуществления подобной реакции и поэтому нужны вещества, которые могут накапливать энергию поглощаемых фотонов до требуемого уровня. В зеленых растениях эту роль выполняет порфирин магния (MgN4C55H72O5), известный как хлорофилл-А, сосредоточенный в хлоропластах растений; он поглощает свет во всем видимом спектре, но особенно сильно в красной области. Отсутствие красного цвета в отраженном растениями свете и дает резко выраженный зеленый цвет. Полагают, что это вещество выполняет каталитическую функцию, переходя в одно из своих триплетных состояний.
Энергия, необходимая для протекания реакции фотосинтеза, велика. С учетом энергии, затрачиваемой на восстановление углекислого газа, она достигает в целом около 5 эВ на единицу синтезируемого углевода. Однако этот процесс может происходить под действием радиации с длинами волн, лежащими за пределами красного конца видимой области спектра, которым соответствуют энергии фотонов лишь около 1,7 эВ. Такие фотоны не учитываются при определении полного к. п. д. процесса, тем не менее их роль в этом процессе пока остается неясной. Большинство исследователей считают, что для синтезирования одной молекулы углевода требуется около восьми фотонов красного света. Если это так, то потребляемая при этом энергия достигает примерно 14 эВ. Поскольку запасенная энергия составляет около 5 эВ, то общий к. п. д. процесса накопления энергии при облучении красным светом равен 35%. Предполагается, что потери возникают на различных стадиях фотосинтеза, большинство из которых принято считать до некоторой степени необратимыми.
В гл. 8 мы видели, как определяется общий к. п. д. использования солнечной энергии в простейшем квантовом процессе. Если порог фотосинтеза соответствует самому краю красного конца спектра (λ ≈ 0,7 мкм), то значительная часть солнечной энергии (около 55%), приходящаяся на инфракрасную область спектра, остается неиспользованной. Более того, если процесс фотосинтеза в чем-то подобен фотоэлектрическому процессу, то и короткие волны в нем не участвуют. Для простоты предположим, что к фотосинтезу применим рассмотренный в гл. 8 метод расчета к. п. д. фотоэлектрического процесса, учитывающий граничную длину волны (там мы получили к. п. д. = 35%). Тогда максимальный к. п. д. использования растениями солнечной энергии оказывается не более.11%. Близкое к этому значение к. п. д. получают при выращивании растений в искусственных условиях, тогда как для естественных условий развития растений к. п. д. на порядок ниже, около 1%. Это и есть предел, к которому стремятся ученые, занимающиеся растениеводством и биологией почвы, и который дает нам надежду, что земля еще сможет нас прокормить.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить ссылку на страницу источник:
http://physiclib.ru/ 'Библиотека по физике'